
ABSTRACT
Areally extensive overturn of deep toxic or biologically unconditioned water, at the beginning of climatic change, is suggested as a possible contributing factor to mass extinction events in the oceans. The overturn and transfer to the photic zone of such waters is due to disruption of the stable stratification of the oceans as the source of oceanic deep water shifts from middle to high latitudes during climatic cooling. In such a weakly stratified ocean, normal geophysical phenomena such as internal tides, heat flow, etc., could provide the uplift. Comparison of extinction events, climate, sulfur-isotope data, pyrite burial rates, sea-level changes, with times of potential overturn since the Late Precambrian suggest that overturn was a contributing factor in extinction events reported for the Late Ordovician, Late Devonian, Late Triassic, and Late Cretaceous; and for marine faunal change in the Late Precambrian and Middle Silurian.
INTRODUCTION
Lipps (1970), Raup and Sepkoski (1982), Sepkoski (1982), Boucot (1983), and McLaren (1983) have discussed extinctions in the marine realm. In particular, Sepkoski (1982) recognized three types of extinctions: one major in the Late Permian; four intermediate in Late Ordovician, Late Devonian, Late Triassic and Late Cretaceous; and several lesser extinctions as reported in the Cambrian (Palmer, 1965; Taylor, 1968; Stritt, 1971), Late Jurassic and early Late Cretaceous (Fischer and Arthur, 1977) and Late Eocene (Tappan and Loeblich, 1971).
The causes of notable extinctions are active areas of debate, with proponents ranging from neo-catastrophists to strict uniformitarians (see Hallam, 1981b, p. 36; Hsu and Drake, 1982; Keith, 1982). Unquestionably, such complex situations as extinctions of many types of organisms at different times would have numerous contributing causes. Each cause would have influence on both extinction and diversity as a function of the living conditions of the organisms, the intensity and duration of the cause, the penchant or preadaptive ability of organisms to exploit the effects of the cause, etc. Our purpose here is not to review or evaluate the various arguments on the cause or causes of marine extinction events, but to suggest contributing factors based on consideration of potential paleo-oceanographic conditions.
OCEANOGRAPHIC FACTORS
Importance of stratification
The present world ocean is strongly density-stratified below the surface mixed layer (Sverdrup et al., 1942). This stratification in middle and low latitudes suppresses vertical motion so that significant upwelling is limited to the surface wind-mixed layer and generally to the upper 50 m. Accordingly, convective overturn common in lakes (Hutchinson, 1957, pp. 437-440) is absent in the modern open ocean.
During glacial--interglacial climates of today, the consequence of having the bulk of deep oceanic water produced by sinking of cold water from high latitudes, overlain by warmer tropical and subtropical surface water produces the present stable density stratification of the world's ocean (Hesselberg and Sverdrup, 1915; Montgomery, 1958). However, through most of the geologic time, the climate has been mild and non-glacial (Frakes, 1979). The Wilde and Berry (1982) model indicates how the oceanic stratification could be destroyed and overturn made possible. Hypothetical water masses for nonglacial ties are plotted on a temperature--salinity--density diagram ( Fig.1). Figure 1 shows that the oceans lose their stratification when high-latitude water, forming at about 5°C, has the same density as middle-latitude highersalinity water. This potential for neutral stability could occur both at the beginning and end of a glacial episode.
Thus, if oceanic stratification was modified so that overturn could occur, then surface conditions would change as a function of the properties of deeper sea water brought to the surface. These changes would offer evolutionary opportunities to some taxa and disadvantages to others, creating a potential for significant extinctions. In particular, changes would be greatest when the difference between surface waters and deep waters is at a maximum.
Present vertical motions
In modern oceanic upwellings, relatively colder, nutrient-rich waters are brought into the surface layers, enhancing productivity and causing plankton blooms. Occasionally blooms become toxic and other organisms (fish) are killed (Dugdale, 1979). Inhibited productivity has been observed during the initial stages of upwelling even with increased nutrients. Barber et al. (1971) attributed this to lack of "conditioning" of the upwelled water, whereby the increased nutrients and trace metals from depth are not biologically available to the primary producers without some chemical modification. Aquaculture experiments using deep nutrient-rich water also show initial inhibition of productivity (Menzel et al., 1963; Barber et al., 1971) indicating that modern deep water is not conditioned. Upwelling in modern anoxic fjords has produced mass mortalities. There, sulphide-bearing water from a subsurface anoxic layer was upwelled into the surface (Richards, 1965, p. 615). Brongersma-Sanders (1957) discussed this toxic effect of sulphides on aerobic marine organisms. In the open ocean, potentially toxic waters were discovered off Peru during a red-tide condition called "Aquaje" (Dugdale et al., 1977). Denitrification (undetectable nitrate-nitrite), hydrogen sulphide and ammonia were discovered below the mixed layer at 150 - 250 m. Such waters are likely to be toxic if introduced rapidly into the photic zone.
We suggest that red tides, toxic fish-killing blooms, and anoxic upwellings found in modern oceans may be small-scale analogs of marine extinction events in which certain populations expand rapidly while others are killed. Extension of this type of upwelling to a larger areal scale and from deeper source depths, as would be the case in an oceanic overturn, might contribute to mass extinctions.
Comparison with other paleo-oceanographic models
Berry and Wilde (1978) proposed a model for the progressive ventilation of the ocean from an initial anoxic to the present well-aerated state. This model assumes a surface-aerated oceanic layer underlain by a residual anoxic layer which was ventilated by glacially driven pulses of cold oxygen-rich deep water. The ventilation of the deep residual anoxic layer occurred presumably by the end of the Paleozoic. A return to anoxicity in deep waters in the post-Paleozoic is indicated if climates remained warm, prohibiting or inhibiting deep ventilation. In particular, in the Cretaceous, the occurrence of deep-oceanic anoxic sediments has been explained by intermittent expansion of the oxygen minimum zone (Schlanger and Jenkyns, 1976; Jenkyns, 1980) or as a result of wide-spread stagnation (Ryan and Cita, 1977). Wilde and Berry (1982), using the Cretaceous as an example, refined their ventilation model to show that variations in climate, with the concomitant shift in the sites of water mass formation, can produce fluctuations between oxic and anoxic conditions.
The "stagnant ocean" model used by Keith (1982, p. 2623) in his explanation of marine extinctions is similar in form to the Wilde and Berry (1982) model used here, with both models generating anoxic conditions below the wind-mixed layer. The "stagnant ocean" implies a strong density stratification in non-glacial times similar to that seen today in the Cariaco trench, Black Sea, and anoxic fjords. In contrast, the Wilde and Berry (1982) model indicates weaker stratification in nonglacial times. Undoubtedly "stagnant ocean" conditions occur locally and could occur locally in the past. The Wilde and Berry "open ocean" concept with conventional water mass formation and mixing (Fig.1) is more applicable to models of global oceanic events. It avoids the biological and mass-balance problems associated with the excess salinity required for strong stratification during mild climates; and provides a mechanism for upwelling toxic water into the photic zone, which is crucial to the extinction arguments. This model also provides a mechanism supporting Lipp's (1970) hypothesis that plankton extinctions are accelerated when thermal stratification is weak.
Conditions for oceanic overturn
Overturn in the oceans could result when vertical motion or buoyancy exceeds and is in the opposite direction to gravity. Thus, review of the stability structure of the oceans and the factors that could produce vertically upward motion are pertinent to assess the paleo-oceanographic potential for overturn.
Static stability in the oceans has been characterized by Hesselberg and Sverdrup (1915) as:
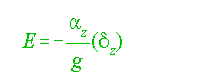 ............................................................................(1)
............................................................................(1)
where:
E = stability;
áz = vertical acceleration;
g = acceleration due to gravity;
z = vertical length.
Otherwise expressed as:
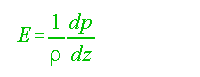 ...........................................................................(2)
...........................................................................(2)
where:
p = density.
For positive E, the water column is stable. For negative E,
the column is unstable and overturn is possible. For E = 0 the column is neutrally
buoyant. Sverdrup et al. (1942, p. 418) used the following approximations.
(1) Above 100 m:
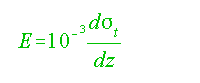 ...........................................................................(3)
...........................................................................(3)
where ót = (P-1) 1000.
(2) Below 100 m, if E by eqn. 3 is less than 40 X 10 - 8then:
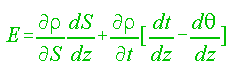 ..........................................................................(4)
..........................................................................(4)
where:
S = salinity;
t = temperature at ambient pressure; and
È= potential
temperature.
As the values of E are on the order of 10-8/ m, a more convenient way to express stability is in terms of the buoyancy frequency (N) (see Gill, 1982, pp. 51--52) where:
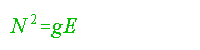 ..........................................................................(5)
..........................................................................(5)
N can also be used to examine dynamic stability via the Richardson number:
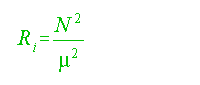
where:
the vertical shear 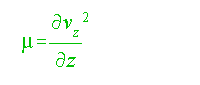 ....................................................................................(6)
....................................................................................(6)
and vz = vertical velocity.
For Ri greater than 0.25, turbulence
will occur (see Munk, 1966, p. 710).
Table I shows static and dynamic stability values of pycnocline conditions for non-glacial to glacial conditions based on tht Wilde and Berry (1982) model (Fig.2). The low values of stability (E and N) and vertical shear (I) in non-glacial times indicate that stratification, although stable, is weak. Consequently, vertical motions and turbulence are easier to initiate than in the strongly stratified glacial ocean.
Unstable water must rise into the photic zone to affect primary productivity and the living conditions of the majority of marine organisms. The potential energy per unit volume required would be:
![]() ...........................................(7)
...........................................(7)
where:
z =
vertical rise; and
As the climate cools, the density of high-latitude water (HLWa) approaches that of middle-latitude water (SMW) (see Fig.1) and the energy per gram for lift decreases drastically. Figure 3 shows the energy per gram contours as a function of climatic cooling at the high-latitude water source and the rise to the photic zone Thus, geophysical phenomena such as tidal shear, planetary waves, internal tides, heat flow, or even micro-seisms might provide sufficient energy to initiate vertical mixing or overturn. These phenomena are of small or local significance in the modern stratified ocean (Munk, 1966). Such forces, however, could be major factors in the overturn of a poorly stratified ocean in advance of actual neutral stratification during climatic transitions.
Heat is transferred faster than salt in the oceans, as the thermal diffusivity is about 100 times the haline diffusivity (Munk, 1966). Consequently, during the time of weak stability at the end of a warm climatic interval, vertical motion might be initiated at the boundary between warmer, saline, bottom water and overlying cooler, fresher, high-latitude water (Fig.1). This situation has been observed in modern oceans as layers in the thermocline above the Mediterranean outflow into the Atlantic (Gregg and Cox, 1972) and in both the Arctic (Neshyba et al., 1971) and the Antarctic (Foster and Carmack, 1976). By analogy with the experiments of Turner (1965) and Griffiths (1979), the water above will separate and rise due to the warming of the fresher layer, producing less dense water. The positive buoyancy of this layer would tend to be maintained or even increased if the separation occurred during the shift in the source of bottom waters from middle-latitude to highlatitude sources. In this manner, the middle-latitude water would further warm the rising boundary layer, increasing its buoyancy. If the oxygen minimum or anoxic water was initially at the boundary, a vertical rise would upwell layers of anoxic or, at least, unconditioned water.
As noted by Riley (1951) and elaborated on by Wyrtki (1962, p. 13), the present rate of oxygen consumption decreases with depth by:
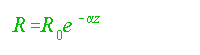 ..................................................(8)
..................................................(8)
where:
R = rate of oxygen consumption;
R0 = R at the surface;
á = property of oxidizable substrates;
z = depth.
For the modern ocean Ro = 1 X 10-8 ml /l per second and á =3.5 X 10-5 cm-1 gives a reasonable fit (Wyrtki, 1962, p. 13). Wyrtki (1962) suggested that most oxidizable material is consumed in the upper layer and or decreases with depth. This relationship would hold if the waters in the ancient pycnocline were oxic. Deep material would be refractory and unconditioned and there would be little immediate effect by contact with surface-ventilated waters during overturn.
However, if anoxic conditions existed below the surface, the value of would not decrease in the anoxic layer as reduced dissolved species would maintain an oxygen demand. In present anoxic fjords, the ratio of H2S to NH3is 3.3 to 1 (Richards, 1965, p. 626). To oxidize the equivalent of one mole of NH3 to NO3- and 3.3 moles of H2S to SO4-, 8.6 moles of O2 are required. Thermodynamically, the sequence of oxidation would be: (1) nitrification (NH3 to NO3-); (2) sulfide oxidation (S2-to SO4-); and (3) aerobic respiration (organic matter to CO2) (Stumm and Morgan, 1970, pp. 336- 337). The actual oxygen consumption, as the deep-water layer rises, would be a function of the oxidation--reduction potential in the layer and the mixing or diffusion at any depth with the ambient oxygen. This can be translated to a redox buffer capacity (Thorstenson, 1970). Brewer and Murray (1973, p. 814) have applied this concept to the oxic/anoxic mixing zone in the Black Sea. Their calculations show that chemosynthetic bacteria use the reduced ionic species. "Bacterial populations at the interface act as a fairly effective lid on the anoxic basin" (Brewer and Murray, 1973, p. 817), preventing the toxic anoxic water from reaching the surface layer. This barrier effect may be the explanation for the existence of anoxic subsurface waters in the Paleozoic, Early Jurassic, and Cretaceous (Wilde and Berry, 1982) without any dramatic evolutionary consequences. Also, modern examples, noted above, of denitrification and the presence of hydrogen sulfide in nearsurface waters (Dugdale et al., 1977) suggest oxidation would not occur if the rise is faster than the modern average vertical rise of 1.2 cm per second (Munk, 1966, p. 719).
Below the mixed layer, reduced conditions would be expected to increase with continued non-glacial climates and more sluggish circulation. Accordingly, an overturn during a climatic transition (Figs.1 and 2, Case A to Case B) after a long non-glacial interval would tend to be more "toxic" to aerobic organisms. At such times both anoxic and unconditioned water would be upwelled
Geographic sequence for overturn
By analogy with the Wilde and Berry (1982) model for the cause of Cretaceous anoxic oceanic events (Arthur, 1979), potential for overturn would begin at middle latitudes. The middle-latitude water (SMW), instead of forming deep water, would either merge with like-density high-latitude water (HLW at 5°C) or seek a neutral density above the higher-density high-latitude water (HLW below 5°C). This would propagate an overturn advancing towards the equator The anoxicity in the near-surface (see Dugdale et al., 1977), and presumably the toxicity of the water in the overturn, would increase equatorward under areas of increased productivity Conceivably, after a long non glacial period, the lack of ventilation at the equator may be sufficient to produce anaerobic conditions there first, prior to overturn. The consequences of overturn at the end of a glacial episode, due to the increased ventilation and circulation during glacial times, would not be as drastic. Overturn would be most toxic in equatorial regions as a consequence of the higher productivity and increased oxygen demand at the equatorial divergence.
Timing sequence for overturn
The duration of the rise is important. Too rapid a rise will produce turbulence and mixing, destroying the positive buoyancy before the deeper water reaches the photic zone. During a slow rise, the non-refractory toxic or inhibitory components might be oxidized or modified by interaction with the surface-ventilated waters.
The sequence of events during an overturn has an influence on the amount of evolutionary opportunity or stress generated by a given climatic event For a relatively slow rate of climatic change, water from depth may be introduced to the photic zone periodically on a seasonal, or other longer-term intervals. Such pulsations would apply pressure to taxa with limited tolerances while permitting adaptable groups to take advantage of the increase in nutrients. Such intermittent anoxicity introduced in the surface layer has been recorded on a small scale for Norwegian fjords, where sulfide-bearing water has produced mass mortality (Richards, 1965, p. 615). If the rate of rise is slow or the stratification is weak so that mixing can produce conditioned nutrient-rich water, overall surface productivity is enhanced. Moreover, the increased productivity would secondarily create an oxygen demand such that anoxicity beneath the surface-ventilated layer could be maintained. Stress on organisms could be prolonged by conventional periodic upwelling from the anoxic layer.
A rapid rate of climatic change causing a single-event overturn may produce mass mortalities in some groups without evolutionary significance, similar to the situation observed in modern fjords. In this case, survivors can recolonize from sanctuaries without having to face continued anoxicity. Accordingly, overturn during a relatively slow rate of climatic change combined with periodic upwellings from a persistent anoxiczone would produce the evolutionary stress needed to explain the extinction of susceptible organisms.
Consequences of overturn on organisms
The Raup and Sepkoski (1982) (Fig.4) curves show that representatives of most families survive extinction events. Thus, any extinction mechanism must allow for survival of, or sanctuaries for, most taxa. The effects on organisms of the overturn described above would be of two types: (1) deprivation or reduction of nutrients or food supplies through inhibition of productivity by aerated but unconditioned waters; followed by (2) direct toxicity of anoxic waters to aerobic organisms. Deleterious conditions must persist at least over the time-span of the maximum breeding/reproductive cycle to insure that surviving life-stages cannot repopulate the threatened environment.
Upwelled anoxic waters not only affect near-surface planktonic and nektonic organisms, but also benthic shelf species. The habitable area required by aerobic bottom dwellers would diminish as anoxic water spread shoreward across the shelf. Benthic species on the outer shelf, would be more likely to experience anoxic or unconditioned waters than inner shelf species, where ventilation to the bottom is assured by wave action and wind mixing. Potentially, inner shelfffea benthic organisms might suffer fewer extinctions than those on the outer shelves.
The marine groups most adversely affected by an overturn would be those with: (1) a limited inhabited area; (2) small population; (3) sessile habit on outer shelf; (4) a benthic life stage, particularly if outer shelf; (5) limited warm temperature tolerance (mainly planktonic); (6) short life-span or reproductive cycle. The marine groups best able to survive an overturn would be those with: (1) widely distributed inhabited area; (2) large population size; (3) tolerance (by encysting, etc.) to anoxic or low-oxygen conditions; (4) high mobility or avoidance behavior; (5) long life or long-term fecundity.
The habitats of extinct organisms and the origin of their replacements as well as the characteristics of survivors would be clues to the efficacy of overturn to contribute to significant extinctions. That examination of the paleontologic record is beyond the scope of this paper.
CORRELATION OF EXTINCTION EVENTS WITH TIMES OF POTENTIAL OVERTURN
Figure 4 shows comparisons among the extinction curves of Raup and Sepkoski (1982) and various phenomena which may reflect oceanic stability and toxicity conditions such as climate, sea-level changes, black-shale episodes, and pyrite burial rates.
Influence of climate and climate-related events
From the foregoing discussion, potential for widespread overturn would occur when the source of deep water shifted from middle to high latitudes (cooling) or from high to middle latitudes (warming). Such conditions always would occur in a climatic transition between non-glacial and true glacial (major sea-ice formation and ice on the continents) conditions Glaciation is not necessary for the density shift, as neutral or near-neutral stratification could occur during fluctuations between warm and cool climates without passing into or out of a glaciation. The conditions for overturn must happen twice per glacial episode as the HLW passes through 5°C (Fig.1); first during a cooling, then during a rewarming after glaciation. Also, overturn could significantly precede or lag behind a glacial event as a function of the rate of cooling or warming. Accordingly, knowledge of the climate and, in particular, the sequence of climatic events is required to estimate the geologic timing of overturns.
The following is a summary of climatic events based chiefly on Frakes (1979). Glacial and cooling intervals from his studies are plotted as bars on the general cooling--warming (evaporite) curve of Meyerhoff (1970) in Fig.4, row II.
Global climates appear to have been relatively warm during most of the Cambrian and Ordovician after episodic glaciation in the Late Precambrian (Frakes, 1979, p. 57). A significant glaciation took place in the latest Ordovician--earliest Silurian (Berry and Boucot, 1973). Frakes (1979, p. 109) concluded that an Early Silurian warming trend was followed by cooling in the Late Silurian into the Middle Devonian. "followed bv a warm Late Devonian". Boucot (1975) noted marked brachiopod faunal provincialism in the late Early--early Middle Devonian The provincialism was influenced by a relatively cool interval in the early Middle Devonian. Frakes (1979, p. 129) pointed out that "widespread glaciations of Gondwanaland characterize the Late Paleozoic interval", and that they range in age from "about Middle Carboniferous into the early Late Permian". Glaciation may have taken place in northeastern Siberia in the Middle and Late Permian (Frakes, 1979). Frakes (1979) indicated that Early Mesozoic climates were cool, but that they warmed during the Triassic. Meyerhoff (1970) (see Fig.4, row II) suggested a cooling trend in the Middle Jurassic. Most of the Jurassic and Cretaceous climates appear to have been warm. Climatic fluctuations have been indicated within the Cretaceous, and climates cooled late in the Cretaceous (Frakes, 1979). Arthur (1979, p. 1478) noted a brief cooling in the Cenomanian--Turonian (about Middle Cretaceous) followed by a brief warming. General cooling followed from the Santonian into the Cenozoic (Arthur, 1979). Cenozoic climates fluctuated markedly (Frakes, 1979). Significant cooling occurred in the Late Eocene into Oligocene and during the Pleistocene glaciations.
Relatively warm intervals during which anoxic waters developed below the wind-aerated surface waters are indicated by widespread, time-synchronous, organic-rich, black shales (Fig.4, row IV). These shales formed during the Late Cambrian--Early Ordovician, Early Silurian, Late Devonian (Berry and Wilde, 1978), Toarcian (Early Jurassic) (Hallam, 1981a), and the Middle Cretaceous deep-sea anoxic events (Jenkyns, 1980).
Brongersma-Sanders (1971) noted the cyclic relationship between evaporites and bituminous rocks Thus, sulfate-isotope (Holler, 1977) and pyriteburial (Berner and Raiswell, 1983) curves (Fig.4, rows III and IV) also may be indicators of general climatic and oceanic ventilation conditions. Sulfatereducing bacteria fractionate sulfur, producing isotopically light sulfides and residual heavy sulfates (see Hoefs, 1980). Accordingly, during times of high pyrite burial (high sulfide production), the sulfate in sea water would be enriched in 34äS, which is reflected geologically in high 34äS values in evaporites. An increasing rate or high values of pyrite burial with high 34äS sulfate values (high evaporation) suggest warming and the potential for widespread development of an anoxic oceanic layer. Decreasing rates or low pyrite burial with lower 34äS sulfate values suggest cooling and ventilation of the ocean.
Potential overturns
The evidence for glaciations and for significant climatic cooling or warming trends summarized in Fig.4 indicates that overturn preceding glaciation could have taken place in the Late Precambrian, Late Ordovician, Middle Carboniferous, and Plio-Pleistocene. If surface water temperatures at high latitudes approached 5°C during cooling trends, then opportunity for overturn existed in the Middle Silurian, Late Triassic, Late Cretaceous, and Late Eocene. Potentially, overturn could take place during warming after a glaciation, as surface waters warm above 5°C in the high latitudes. Such conditions could have occurred in the Latest Precambrian, Middle to Late Devonian, Late Permian, and latest Pleistocene--Recent. If a residual oceanic anoxic layer, as proposed by Berry and Wilde (1978), survived the glacial ventilation, a postglacial overturn may be as deleterious as a preglacial overturn. This potential probably vanished with the ventilation of the oceans during the great Permo-Carboniferous glaciation. Since then, with increased ventilation and oceanic circulation during cold or cooler periods, the effect on biota of an overturn after the cold period would be less In any case, the potential for mass extinctions would be greatest during overturn after a prolonged warm period because of the inherent increased anoxicity due to sluggish oceanic circulation.
Influence of sea level and shelf area fluctuations
Hallam (1981b) noted that living area as well as climatic change may have an effect on speciation and extinction rates among marine organisms. Newell (1967), Schopf (1974) and Simberloff (1974) have discussed the effects of reduction of habitable space on marine benthic organisms citing especially the Late Permian when marine invertebrate extinctions increased as habitable area on the shelf decreased. In general, transgressions to higher sea levels implies increased shelf seas, whereas regressions to lower sea levels would reduce marine shelf areas. Sea-level changes with time (Shanmugan and Moila, 1982) are shown in Fig.4, row V. However, comparison of the sea-level curve with the Raup and Sepkoski (1982) data (Fig.4) reveals that sea-level changes and glaciations alone had seemingly little effect on marine life. For example, major regressions associated with Carboniferous and Pleistocene glaciations are not reflected as times of major marine extinctions. Certain extinctions might be related to transgressions. In such cases the area of inhabitable shelf could be reduced during transgressions, even though the area of the shelf sea would be increased, by the spread of anoxic waters from the oceanic anoxic layer onto the shelf (Hallam, 1981a, p. 225). This would appear as a zone below the surface wind-mixed layer approximately 100 m below the absolute sea level (see Berry and Wilde, 1978). The potential anoxic zone is shown on Fig.4, row V as between the absolute depth of the edge of the shelf ( - 200 m) and within 100 m of the sea surface. Whether anoxic bottom water is in this zone is a function of the ventilation of the ocean at that time. Potentially, the most deleterious situation for shelf organisms may occur at high stands of sea level when broad expanses of the shelf floor are covered with anoxic water. At such times, episodic introduction of anoxic waters into the nearshore and surface-ventilated waters is more likely. The combined factors of reduced living space, widespread climatic change, and spread of toxic waters into shallow marine environments appear to have influenced extinctions or contributed to significant changes among marine organisms.
SUMMARY AND CONCLUSIONS
The potential for oceanic overturn may occur when the source for deep water shifts from middle to high latitudes (a cooling trend) or from high to middle latitudes (a warming trend) (see Fig.1 and Fig. 2). If anoxic waters had developed widely throughout the oceans prior to overturn, then the possibilities existed for widespread marine extinctions.
Overturn may occur as oceanic middle waters become relatively neutrally buoyant in relation to deep waters. Under near-neutral buoyancy, a number of influences, which include heat flow and internal waves, that today are relatively insignificant to present water-column stability, may induce overturn. If overturn is either relatively rapid or very slow, then a rise of toxic waters may have little effect on organisms. Under appropriate rates of upwelling, toxic waters may reach the surface or near-surface and spread across the normally ventilated parts of continental shelves. As toxic waters spread, potential for extinctions and rapid faunal turnover is enhanced.
The lack of correlation among major marine extinctions (see Fig.4, rows I and V) with sea level fall and onset of cold climates, such as those of the Middle Carboniferous, Middle Tertiary, and Pleistocene, indicates that these factors alone do not result in extinctions. This may be the result of insufficient time since the last cooling to develop toxic or unconditioned water to be upwelled during an overturn.
Certain other faunal changes noted in the marine record [including Sepkoski's (1982) "lesser extinctions"] may be reflective of oceanic overturn. For example, loss of the Vendian faunal elements (Raaben, 1981) in the Late Precambrian may be related to overturn following climatic warming. An oceanic overturn could explain the rapid turnover in both early and late Middle Silurian graptolites (see Rickards et al., 1977) and the turnover in Early Silurian shelf marine benthic communities (Boucot, 1983). Overturn coupled with climatic cooling and a reduced area of shelf seas could account for the Late Triassic extinctions (Hallam, 1981b).
In certain marine extinctions, (such as the Late Ordovician, Late Cretaceous, and possibly Late Devonian) a significant change occurs among planktonic or nektonic organisms living over outer shelves and bathyal environments as well as benthic organisms living primarily in outer-shelf environments. In such cases, oceanic overturn, in which toxic waters are upwelled into these environments, may be a potential cause for extinctions and rapid faunal turnovers. Table II summarizes our views on which potential overturns contributed to extinction and faunal changes.
ACKNOWLEDGEMENTS
We are grateful to A. T. Dengler for his technical review, particularly with respect to the physical oceanographic model and computations. Useful discussions with R. A. Berner and students at a seminar given on this work at Yale University were greatly appreciated. Word Processing was handled by J. Edgar and the illustrations were designed and executed by M. Arcidiacono Krup. The research was supported in part by the U.S. Department of Energy (contract DE-AT03-83CE89302).
REFERENCES
Arthur, M. A., 1979. Paleoceanographic events--recognition, resolution, and reconsideration. Rev. Geophys. Space Phys., 17: 1474 - 1494.
Barber, R. T., Dugdale, R. C., MacIsaac, J. J. and Smith, R. L., 1971. Variations in phytoplankton growth associated with the source and conditioning of upwelling water. Invest. Pesq., 35: 171--193.
Berner, R. A. and Raiswell, R., 1983. Burial of organic carbon and pyrite sulfur in sediments over Phanerozoic time: a new theory. Geochim. Cosmochim. Acta, 47: 855-- 862.
Berry, W. B. N. and Boucot, A. J., 1973. Glacio-eustatic control of Late Ordovician--Early Silurian platform sedimentation and faunal changes. Geol. Soc. Am. Bull., 84: 275-- 284.
Berry, W. B. N. and Wilde, P., 1978. Progressive ventilation of the oceans--an explanation for the distribution of the Lower Paleozoic black shales. Am. J. Sci., 278: 257 - 275.
Boucot, A. J., 1975. Evolution and Extinction Rate Controls. Elsevier, Amsterdam, 427 pp.
Boucot, A. J., 1983. Does evolution take place in an ecological vacuum? II. J. Palaeontol., 57: 1 - 30.
Brass, G. W., Southam, J. R. and Peterson, W. H., 1982. Warm saline waters in the ancient oceans. Nature, 295: 620--623.
Brewer, P. G. and Murray, J. W., 1973. Carbon, nitrogen and phosphorus in the Black Sea. Deep-Sea Res., 20: 803 - 818.
Brongersma-Sanders, M., 1957. Mass mortality in the sea. In: J. W. Hedgpeth (Editor), Treatise on Marine Ecology and Paleoecology. Geol. Soc. Am. Mem., 67(1): 941--1010.
Brongersma-Sanders, M., 1971. Origin of major cyclicity of evaporites and bituminous rocks: An actualistic model. Mar. Geol., 11: 123 - 144.
Chamberlain, T. C., 1906. On a possible reversal of deep-sea circulation and its influence on geologic climates. J. Geol., 14: 371 - 372.
Dugdale, R. C., 1979. Primary nutrients and red tides in upwelling regions. In: D. L. Taylor and H. H. Seliger (Editors), Toxic Dinoflagellate Blooms. Elsevier, Amsterdam, pp. 257 - 262.
Dugdale, R. C., Goering, J. J., Barber, R. J., Smith, R. L. and Packard, T. T., 1977. Denitrification and hydrogen sulfide in the Peru upwelling region during 1976. DeepSea Res., 24: 601 - 608.
Fischer, A. and Arthur, M. A., 1977. Secular variations in the pelagic realm. Soc. Econ. Paleontol. Min., Spec. Publ., 25: 19--50.
Foster, T. D. and Carmack, E. C., 1976. Temperature and salinity structure in the Weddell Sea. J. Phys. Oceanogr., 6: 36--44.
Frakes, L. A., 1979. Climates Throughout Geologic Time. Elsevier, Amsterdam, 310 pp.
Gill, A. E., 1982. Atmosphere--Ocean Dynamics. Academic Press, New York, NY,662 pp.
Gregg, M. C. and Cox, C. S., 1972. The vertical microstructure of temperature and salinity. Deep-Sea Res., 19: 355 - 376.
Griffiths, R. W., 1979. The transport of multiple components through thermohaline diffusive interfaces. Deep-Sea Res., 26A: 383 - 397.
Hallam, A., 1977. Jurassic bivalve biogeography. Paleobiology,3: 58 - 73.
Hallam, A., 1981a. Facies Interpretation and the Stratigraphic Record. W. H. Freeman, San Francisco, CA, 291 pp.
Hallam, A., 1981b. The end-Triassic bivalve extinction event. Palaeogeogr., Palaeoclimatol., Palaeoecol., 35: 1--44.
Hess, H. H., 1962. History of ocean basins. In: A. E. J. Engel, H. L. James and B. F. Leonard (Editors), Petrologic Studies: A Volume in Honor of A. F. Buddington. Geol. Soc. Am., pp. 599--620.
Hesselberg, T. and Sverdrup, H. V., 1915. Die Stabilitats-Verhaltnisse des Seewassers bei vertikalen Verschiebungen. Berg. Mus. Aarb., 15: 16 pp.
Hoefs, J., 1980. Stable Isotope Geochemistry. Springer-Verlag, Heidelberg, 203 pp.
Holser, W. T., 1977. Catastrophic chemical events in the history of the ocean. Nature, 267: 403 - 408.
Hsu, K. and Drake, C. L., 1982. Comment and reply on "Impacts and Evolution Conference". Geology, 10: 557--558.
Hutchinson, G. E., 1957. Treatise on Limnology. Wiley, New York, NY, 1017 pp.
Jablonski, D. and Bottjer, D. J., 1981. Soft-bottom epifaunal suspension-feeding assemblages in the Late Cretaceous: implications for the evolution of benthic communities. Geol. Soc. Am., Abstr. Progr., 13: 7: 479.
Jenkyns, H. C., 1980. Cretaceous anoxic events: from oceans to continents. Geol. Soc. Lond. J., 137: 171--188.
Keith, M. L., 1982. Violent volcanism, stagnant oceans and some inferences regarding petroleum, strata-bound ores and mass extinctions. Geochim. Cosmochim. Acta, 46: 2621 - 2637.
Lipps, J. H., 1970. Plankton evolution. Evolution, 24; 1 - 22.
McKerrow, W. S., 1978. The Ecology of Fossils. The MIT Press, Cambridge, MA, 384 pp.
McLaren, D. J., 1983. Bolides and biostratigraphy. Geol. Soc. Am. Bu11.,94: 313--324.
Menzel, D. W., Hulburt, E. M. and Ryther, J. H., 1963. The effects of enriching Sargasso Sea water on the production and species composition of the phytoplankton. Deep-Sea Res., 10: 209--219.
Meyerhoff, A. A., 1970. Continental drift, II. High-latitude evaporite deposits and geologic history of Arctic and North Atlantic Oceans. J. Geol., 78: 406 - 444.
Monin, A. S., Kamenkovich, V. M. and Kort, V. G., 1977. Variability of the Oceans. Wiley, New York, NY, 241 pp. (translated from Russian).
Montgomery, R. B., 1958. Water characteristics of Atlantic Ocean and World Ocean. Deep-Sea Res., 5: 134--148.
Munk, W. H., 1966. Abyssal Recipes. Deep-Sea Res., 13: 701--730.
Neshyba, S., Neal, V. T. and Denner, W., 1971. Temperature and conductivity measurements under Ice Island T-3. J. Geophys. Res., 76: 8107 - 8120.
Newell, N. D., 1967. Revolutions in the history of life. Geol. Soc. Am., Spec. Pap., 89: 63 - 91.
Palmer, A. R., 1965. Biomere--a new kind of biostratigraphic unit. J. Paleontol., 39: 149 - 153.
Raaben, M. E. (Editor), 1981. The Tommotian Stage and the Cambrian lower boundary problem. U.S. Department of the Interior and the National Science Foundation Washington, DC, 359 pp. (translated from Russian).
Raup, D. M. and Sepkoski, J. J., 1982. Mass extinctions in the marine fossil record. Science, 215: 1501 - 1503.
Reid, J. L., 1965. Intermediate Waters of the Pacific Ocean. Johns Hopkins Press, Baltimore, MD, 85 pp.
Richards, F. A., 1965. Anoxic basins and fjords. In: J. P. Riley and G. Skirrow (Editors) Chemical Oceanography, 1, Academic Press, New York, NY, pp. 611--645.
Rickards, R. B., Hutt, J. E. and Berry, W. B. N., 1977. Evolution of the Silurian and Devonian graptoloids. Br. Mus. (Nat. Hist.), Geol., 28: 120 pp.
Riley, G. A., 1951. Oxygen, phosphate, and nitrate in the Atlantic Ocean. Yale Univ. Bull. gingham Oceanogr. Coll., 13: 1 - 126.
Ryan, W. B. F. and Cita, M. B., 1977. Ignorance concerning episodes of world-wide stagnation. Mar. Geol., 23: 197 - 215.
Schlanger, S. O. and Jenkyns, H. C., 1976. Cretaceous oceanic anoxic events: causes and consequences. Geol. Mijnb., 55: 179 - 184.
Schopf, T. J. M., 1974. Permo-Triassic extinctions: relation to sea-floor spreading. J. Geol., 82: 129 - 143.
Sepkoski, J. J., 1982. Mass extinctions in the Phanerozoic oceans: a review. Geol. Soc. Am., Spec. Pap., 190: 281--289.
Shanmugan, G. and Moila, R. J., 1982. Eustatic control of turbidites and winnowed turbidites. Geology, 10: 231--235.
Simberloff, D., 1974. Permo-Triassic extinctions: effects of area of biotic equilibrium. J. Geol., 82: 267--274
Stitt, J. H., 1971. Repeating evolutionary pattern in Late Cambrian Trilobite Biomeres. J. Paleontol., 45: 178--181.
Stumm, W. and Morgan, J. J., 1970. Aquatic chemistry, Wiley-Interscience, New York, NY, 583 pp.
Sverdrup, H. V., Johnson, M. A. and Fleming, R. A., 1942. The Oceans. Prentice-Hall, Englewood Cliffs, NJ, 1042 pp.
Tappan, H. and Loeblich, A. R., 1971. Geologic implications of fossil phytoplankton evolution and time--space distribution. Geol. Soc. Am, Spec. Pap., 127: 247 pp.
Taylor, M. E., 1968. The Mindyallan fauna of North-Western Queensland, by A. A. Opik (review). J. Paleontol., 42: 1319--1321.
Thorstenson, D. C., 1970. Equilibrium distribution of small organic molecules in natural waters. Geochim. Cosmochim. Acta 34: 745--770
Turner, J. S., 1965. The coupled transports of salt and heat across a sharp density interface. Int. J. Heat Transfer, 8: 759--767.
Vail, P. R. Mitchum, R. M., Jr. and Thompson, S., III, 1977. Seismic stratigraphy and global changes of sea level, part 4: Global cycles of relative changes of sea level. In: D. J. Stanley and G. Kelling (Editors), Seismic Stratigraphy--Applications to Hydrocarbon Exploration. Am. Assoc. Pet. Geol. Mem., 26: 83--97.
Wise, D. U., 1972. Freeboard of the continents through time. In: Studies in Earth and Space Sciences. Geol. Soc. Am. Mem., 132: 87--100.
Wilde, P. and Berry, W. B. N., 1982. Progressive ventilation of the oceans--potential for return to anoxic conditions in the post-Paleozoic. In: S. O. Schlanger and M. B. Cita (Editors), Nature and Origin of Cretaceous Carbon-Rich Facies. Academic Press, New York, NY, pp. 209--224.
Worthington, L.V., 1959. The 18°C water in the Sargasso Sea. Deep-Sea Res., 5: 297-- 305.
Wyrtki, K., 1962. The oxygen minimum in relation to ocean circulation. Deep-Sea Res., 9: 11--23
TABLE I
| Units | Climate | ||
|---|---|---|---|
| Non-glacial | Pre- or Post glacial |
Glacial (Modern) | |
| 108 E (per meter) | 43 | 84.0 | 523 |
| 104N/2 Ð (Hertz) | 3 | 4.5 | 11 |
| 2 Ð/N X 60 (minutes) | 51 | 36.8 | 14.8 |
| 104µ vertical (per second) | 6 | 8.9 | 21 |
TABLE II
| Time of overturn (Fig.4. row H) | Type | Prior black shale (Fig.4, row IV) | Anoxic shelf (Fig.4. row V) |
Extinction events (Fig.4, row I) | Overturn as Factor |
|---|---|---|---|---|---|
| Late Precambrian | Preglacial Postglacial | ? ? | Likely Likely |
Not Reported Vendian faunal change (Raaben, 1981) | ? Yes |
| Late Ordovician | Preglacial | Yes | Likely | Raup and Sepkoski (1982) (event 1) | Yes |
| Middle Silurian | Postglacial | Likely | Faunal change, graptolites (Rickards et al., 1977); benthic (McKerrow 1979) | Yes | |
| Mid-Late Devonian | Cooling | Yes | Likely | Raup and Sepkoski (1982) (event 2) | Yes |
| Middle Carboniferous | Preglacial | No | Possible | Not reported | |
| Late Permian | Postglacial | No | Unlikely | Raup and Sepkoski (1982) (event 3) | Unconditioned water only |
| Late Triassic | Cooling | No | Possible | Hallam (1981b) (event 4) | Yes |
| Late Cretaceous | Cooling | Deep-sea Yes |
Possible | Fischer and Arthur (1977) Raup and Sepkoski (1982) (event 5) | Yes |
| Late Eocene | Cooling | No | Unlikely | Tappan and Loeblich <(1971) | unconditioned water only |
| Plio-Pleistocene | Preglacial | No | unlikely | not reported |